Chemotaxis in rhodobacter sphaeroides: a feasibility study

Chemotaxis in Rhodobacter sphaeroides
A Feasibility Study
Project Members
- Antonis Papachristodoulou
- Mark Roberts
- Elias August
- Abdullah Hamadeh
- Nassia Inglessis
Introduction
Recent years have seen the application of mathematical modelling to an increasing number of problems in the biological and medical sciences. A mathematical model can represent an ecosystem, an organ, a cell, or the kinetic reaction between molecules. It can provide useful insights into the dynamics of biological systems and, importantly, guide the scientists who might be overwhelmed by the complexity of their research objects. Many biological processes involve crosstalk and feed-back loops generating complex networks rather than simple linear pathways. Obtaining the topological structure of such networks is important for understanding the mechanism through which robust system functionally is maintained. High throughout experiments now provide a wealth of data that can be used to determine biochemical network structure and to propose mechanistic rate laws with appropriate kinetic parameter values. However, no unique network can account for these data, and the systematic design of new experiments based on current knowledge is essential for further delineating network structure. A synergy between mathematical modelling, control theory and experiment design is therefore fundamental for increasing physiological and biochemical knowledge.
Bacterial chemotaxis is the biasing of movement towards regions of higher concentration of beneficial or lower concentration of toxic chemicals. In bacteria such as Escherichia coli and Rhodobacter sphaeroides, this is achieved when chemical ligands bound to membrane-spanning receptors initiate a signalling cascade of intracellular protein activity leading to the change in activity of the flagellar motor which drives the extracellular flagellum (or flagella), causing the bacterium to move. Chemotaxis in Escherichia coli is one of the best understood pathways in biology and there is a large amount of experimental data on structures, kinetics, in vivo protein concentrations and localisation. This relatively simple pathway has helped to conceptualise the signalling pathway of sensory systems in general. However, with an increasing number of sequenced bacterial genomes it becomes evident that the chemotactical sensory mechanism of many other bacteria is much more complex (Wadhams et al, 2004).
We are developing a theoretical framework in order to design, by judiciously choosing experimental conditions and stimuli, new experiments to differentiate between possible model alternatives. One problem is to find the best initial condition or parameters that maximise the difference between the different models. It can be approached through the notion of observability for a linear system or of a storage function in the case of a nonlinear system. Particularly, for nonlinear systems new and powerful computational tools have been developed to search for this function; one is SOSTOOLS, a recently developed MATLAB toolbox that connects semidefinite programming with the sum of square decomposition. We use these tools also to address the problem of designing the best, possibly time varying, input or pathway deletion that will maximise the differentiability between competing models. In summary, this work intends to optimise the design of experiments that in/validate models and to close the loop between modelling and experiment design to increase our understanding of the connectivity and function of complex biochemical networks, from which the biosciences will benefit greatly. There is a lot to be gained through interdisciplinary research involving scientists from different fields and by borrowing methods from other fields that have a long tradition of working with models of dynamical systems, such as control theory.
This research is supported by EPSRC project E05708X.
Chemotaxis
Bacterial chemotaxis is the biasing of bacterial movement towards regions of higher concentration of beneficial, or lower concentration of toxic, chemicals. This is achieved when chemical ligands bind to membrane-spanning receptors and initiate a signalling cascade of intracellular protein activity leading to the change in the activity of the flagellar motor and causing the bacterium to change the direction in which it moves. This results in a biased random walk (Figure 1).
Chemotaxis in E. Coli
Chemotaxis in E. coli is one of the best understood pathways in biology. This relatively simple pathway has helped to conceptualise the signalling pathway of general sensory systems. However, with an increasing number of sequenced bacterial genomes it becomes clear that the chemotactic sensory mechanism of other bacteria is much more complex.
The signalling pathway within E. coli is well understood and is a simple circuit with one feedback loop (see Figure 2). The receptor in the system is a Methyl-accepting Chemotaxis Protein (MCP) that senses ligands outside the cell. Associated with the MCP is the histidine protein kinase called CheA. Binding of certain ligands to MCP decreases the auto-phosphorylation rate of CheA. CheA can transfer phosphoryl groups to two possible response regulators CheY and CheB. CheY-P interacts with motor binding sites of the multiple E. coli flagella motors causing a change in direction. The receptors are constantly methylated by the action of a methyltransferase CheR, while CheB-P acts as a methylesterase to demethylate the receptor, making it less responsive to ligand binding. This creates a feedback loop, allowing for adaptation. Adaptation allows E. coli to react to changes of the concentration gradient of ligands and not to changes in concentrations per se. Finally, CheZ acts to dephosphorylate CheY and to terminate the signal.
Chemotaxis in R. sphaeroides
The bacterium R. sphaeroides has three chemotaxis operons, of which two are expressed under normal laboratory conditions. Proteins expressed from these operons have previously been shown to localise to discrete signalling clusters, one at the poles of the cell, similar to E. coli, and one in the cytoplasm. This localisation is thought to prevent cross talk between the two clusters, allowing them to potentially signal separately.
In this bacterium, external ligands reduce the number of active receptors and this causes a decrease in the autophosphorylation rate of the CheA homologues, CheA2 and the CheA3A4 complex. Phosphotransfer experiments show that CheA-P homologues can phosphorylate the CheY and CheB homologues: CheA2-P phosphorylates the proteins CheY3, CheY4, CheY6, CheB1 and CheB2, while CheA3A4-P can only phosphorylate CheY6 and CheB2 (Figure 3). When the concentration of phosphorylated CheA2 and CheA3A4 is reduced, the concentrations of phosphorylated CheY3, CheY4, CheY6, CheB1 and CheB2 are also reduced. In the forward signalling pathway, it is suspected that the phosphorylated CheY homologues have a similar role to CheY in E. coli, interacting with the flagellar motors to increase their rate of switching. When the amount of phosphorylated CheY homologues decreases therefore, we can expect the rate of flagellum switching to decrease, thus leading to a bias in the bacterium's random walk. In the feedback pathway, it is suspected that CheB1 and CheB2 have a similar role to CheB in E. coli, acting to demethylate active receptors. Thus, when the concentration of phosphorylated CheB1 and CheB2 is reduced the receptor demethylation rate decreases, resulting in an increase in the number of active receptors. Through this adaptation mechanism, the concentration of active receptors is restored to its basal level prior to the ligand stimulus and the biasing of the bacterium's random walk is removed.
Methodology
Engineering Control Theory
Our method involves the creation of models with different connectivities that can explain current experimental data. The models have in common all currently known connectivities and differ in that each model represents a new speculative connectivity. Given that all models can represent wild type data, we must perturb the system in order to create conditions that allow for the invalidation of some of the models. We do this in two ways (see references Papachristodoulou, 2007: 2714-2719 and Papachristodoulou, 2007: 2872-2877):
Input Design: We mathematically determine the frequency of a sinusoidal input to the system which results in the largest difference between the outputs of the models under consideration using a Bode plot (Figure 4); this is a tool often used in control and systems theory. Thus, if one of the models resonates at a certain frequency while the others do not, then exciting the real system at this frequency will help us to distinguish between the models.
Initial Condition Design: We can also make changes to the initial conditions and then test these changes in silico in order to determine those which will discriminate best between the models under test (Figure 5), before committing resources to undertake in vivo and in vitro experiments. The exact nature of the perturbation that can be performed will vary with the system being investigated, but could include altering protein levels by knockout, knockdown (eg RNAi in eukaryotes), protein over expression, etc.
Biochemistry
Biochemical kinetic constants used for the model creation are being determined using phosphortransfer assays (Porter et al, 2002).
The proposed interconnectivity is also being tested using in vitro SPR of purified components (Biacore) and in vivo two hybrid experiments.
We are employing a tethered cell assay (Figure 6), which allows us to measure the response of the bacterium to controlled external stimuli. This allows us to measure the repsonse of the pathway to a variety of stimuli and correlate this with the modelling.
Elucidating Chemotaxis Signalling Pathways
We take piecemeal approach to elucidating network structure, considering first the forward path from the ligand detection to motor activation and then the feedback path responsible for exact adaptation.
Determining the forward signalling pathway
The R. sphaeroides chemotaxis forward signalling pathway conveys signals arising from sensed ligands to the flagellar motors. We consider four possible forward path models, each of which reproduce ligand response data. The basic characteristics of these models are summarised as follows:
Model A: CheY6 and CheY3 or CheY4 bind to the motor co-operatively
Model B: where CheY3 and CheY4 interact with the Tlps of the central cluster and CheY6 interacts with the flagellar motors.
Model C: where CheY3 and CheY4 interact with the Tlps of the central cluster
Model D: CheY3 and CheY4 acting only as phosphate sinks, similar to the roles of the multiple CheY’s
Using the methodology described above we were able to invalidate all but Model B.
Determining the feedback pathway
In the feedback pathway the proteins CheB1 and CheB2 are responsible for adaptation, acting to demethylate active receptors and thus restoring the chemotaxis system to its pre-stimulus equilibrium level. We can conceive of four different feedback models, each capable of reproducing ligand response data. These models are characterised thus:
Model 1: CheB1-P de-methylates at the transmembrane receptor cluster, and CheB2-P at the cytoplasmic cluster only (shown in solid lines in Figure 7).
Model 2: CheB1-P and CheB2-P both de-methylate at both clusters (solid lines, the dashed line and the dashed-dotted line).
Model 3: CheB1-P and CheB2-P both de-methylate at the membrane cluster and CheB2-P at the cytoplasmic cluster only (solid lines and the dashed line).
Model 4: CheB1-P de-methylates at the membrane cluster, CheB1-P and CheB2-P both de-methylate at the cytoplasmic cluster (solid lines and the dashed-dotted line).
Chemotaxis in R. sphaeroides and the cascade control design principle
The remaining chemotaxis pathway for R. sphaeroides we were unable to invalidate, incorporating both the forward and feedback signalling paths given in Figure 8.
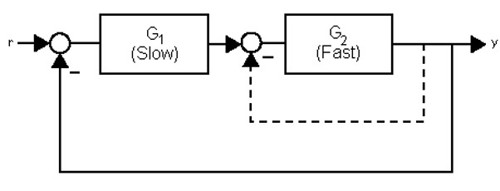
Model 3 has close and interesting links with a control engineering feedback design principle called cascade control. This term denotes the CheB2-P feedback around the fast cytoplasmic cluster embedded within the CheB2-P feedback de-methylating the slow transmembrane cluster.
Figure 9 shows a typical cascade controlled system compared to the R. sphaeroides chemotaxis system (Figure 10). The cascade control architecture improves the performance of systems by increasing their damping and rejecting output disturbances.
Final Project Report
We have met all the targets outlined in the original proposal. We developed a modelling strategy in order to understand the structure of signal transduction networks, iterating between network determination, experiment design and model invalidation. The idea behind this approach is that many mathematical models can explain experimental data for a biological system, but at most one is correct. In order to differentiate between models, one needs to invalidate incorrect models, using appropriate experimental evidence. The aim of this project was to develop a methodological framework to design experiments that would allow invalidation. The methodology that we developed was tested using the phosphotransfer network responsible for chemotaxis in a model organism, Rhodobacter sphaeroides.
As part of this project, we developed three control engineering inspired systems biology tools:
(1) A network determination technique that uses linear programming for efficient, sparse network determination. This technique was applied to different types of networks (transcriptional, metabolic and chemical) using real experimental data - for more details see “Efficient, sparse biological network determination” by E. August and A. Papachristodoulou, BMC Systems Biology 2009, 3:25.
(2) Experiment design algorithms for discrimination, based on initial condition design, dynamic input design and design of structural modifications.
(3) Model invalidation tools in discrete and continuous time, that use experimental data to show that models are invalid - for more details, see “On Validation and Invalidation of Biological Models” by J. Anderson and A. Papachristodoulou, BMC Bioinformatics 2009, 10:132.
Blending the above three layers, we developed an iterative method that involves network determination, experiment design and model invalidation, and applied it successfully to the forward signalling part of the chemotaxis pathway of our model organism, Rhodobacter sphaeroides. We proposed four competing models of the possible signalling mechanism and then designed experiments to invalidate three of them - for more details, see “A model invalidation-based approach for elucidating biological signalling pathways, applied to the chemotaxis pathway in R. sphaeroides” by M. A. J. Roberts, E. August, A. Hamadeh, P. K. Maini, P. E. McSharry, J. P. Armitage and A. Papachristodoulou, BMC Systems Biology 2009, 3:105.
We also repeated our methodology and applied it on the feedback mechanism. Additional research was performed on the identifiability of parameters in biochemical networks from input-output data - for more details, see “A new computational tool for establishing model parameter identifiability” by E. August and A. Papachristodoulou, Journal of Computational Biology. June 2009, 16(6): 875-885. Some preliminary work has been done on the analysis of biochemical reaction networks resulting from this model development cycle.
In addition to the four peer reviewed papers, the work was disseminated at several conferences (IEEE Conference on Decision and Control 2008, International Conference on Systems Biology 2008 and 2009, BioSysBio 2008 and Bacterial Locomotion and Signal Transduction 2009) as well as in several workshops (Wellcome Trust workshop on Engineering Principles in Biology, Weismann Institute on Systems Biology of Cancer, and at two workshops of the Institute of Mathematics and its Applications at the University of Minneapolis: “Organisation of Biological Networks” and “Design Principles in Biological Systems”) and University seminars (Imperial College, University of Manchester, University of Liege, California Institute of Technology).

Figure 1: A – A random walk. B – Chemotaxis biasing a random walk.

Figure 2: The E. coli chemotaxis pathway.

Figure 3: The internal R. sphaeroides phosphorylation cascade.

Figure 4: Designing the ligand input to maximise the output difference between possible chemotaxis pathway models.

Figure 5: Designing the initial conditions to maximise the output difference between possible chemotaxis pathway models.

Figure 6: Schematic view of a tethered cell.

Figure 7: Different possible chemotaxis feedback pathways in R. sphaeroides.

{kind=link}
{kind=link}
{kind=link}

Figure 10: The suggested R. sphaeroides chemotaxis pathway.
Related SySOS Project Publications
Journal Publications
- A. Hamadeh, M. A. J. Roberts, E. August, P. E. McSharry, P. K. Maini, J. P. Armitage and A. Papachristodoulou. Feedback Control Architecture and the Bacterial Chemotaxis Network. PLoS Computational Biology, 7(5): e1001130, 2011. [1]
- M. A. J. Roberts, E. August, A. Hamadeh, P. K. Maini, P. E. McSharry, J. P. Armitage and A. Papachristodoulou. A model invalidation-based approach for elucidating biological signalling pathways, applied to the chemotaxis pathway in R. sphaeroides. BMC Systems Biology 2009, 3:105. [2]
- J. Anderson and A. Papachristodoulou. On Validation and Invalidation of Biological Models. BMC Bioinformatics 2009, 10:132. [3]
- E. August and A. Papachristodoulou. Efficient, sparse biological network determination. BMC Systems Biology 2009, 3:25. [4]
- E. August and A. Papachristodoulou. A new computational tool for establishing model parameter identifiability. Journal of Computational Biology. June 2009, 16(6): 875-885.[5]
Conference Publications
- A. Hamadeh, E. August, M. A. J. Roberts, P. K. Maini, J. P. Armitage, B. P. Ingalls and A. Papachristodoulou. Feedback Control Architecture of the R. sphaeroides Chemotaxis Network. In Proc of the IEEE CDC 2011.
- E. August, A. Papachristodoulou, B. Recht, M. A. J. Roberts and A. Jadbabaie. Determining Interconnections in Biochemical Networks Using Linear Programming. In Proc. of the IEEE CDC, 2008 [6]
References
- Porter, S.L., and Armitage, J.P. (2002) "Phosphotransfer in Rhodobacter sphaeroides Chemotaxis." J Mol Biol 324: 35-45.
- Wadhams, G.H., and Armitage, J.P. (2004) "Making sense of it all: bacterial chemotaxis." Nat Rev Mol Cell Biol 5: 1024-1037.
- Papachristodoulou, A., and El-Samad, H. (2007) "Algorithms for Discriminating Between Biochemical Reaction Network Models: Towards Systematic Experimental Design." Proceedings of the 2007 American Control Conference: 2714-2719
- Papachristodoulou A., and Recht, B. (2007) "Determining Interconnections in Chemical Reaction Networks." Proceedings of the 2007 American Control Conference: 2872-2877